SpellChecker.net
Spellcheck dictionary
Misspelling dictionary
Spellcheck
Misspellings
Basic Check
Advanced Check
Austria - German
Belarus - Belarusian
Catalan - Valencian
Chinese
Danish - Dansk
Dutch - Nederlands
English - Australia
English - Canada
English - United Kingdom
English - United States
English - GB
English - New Zealand
Esperanto
France - Breton
French - Français
German
German - Deutsch
German - Simple
German Switzerland - Schweizerdeutsch
Greek - Ελληνικά
Iran - Persian
Italian - Italiano
Japanese
Khmer
Philippines - Tagalog
Polish - Polski
Portuguese
Portuguese - Português
Portuguese Brazil - Português
Portuguese - Angola
Portuguese - Moçambique
Romanian - Română
Russian - Русский
Slovak - Slovenčina
Slovenian - Slovenščina
South Africa - English
Spanish - Español
Spain - Asturian
Spain - Catalan
Spain - Galician
Swedish - Svenska
Tamil
Ukrainian - Українська
×
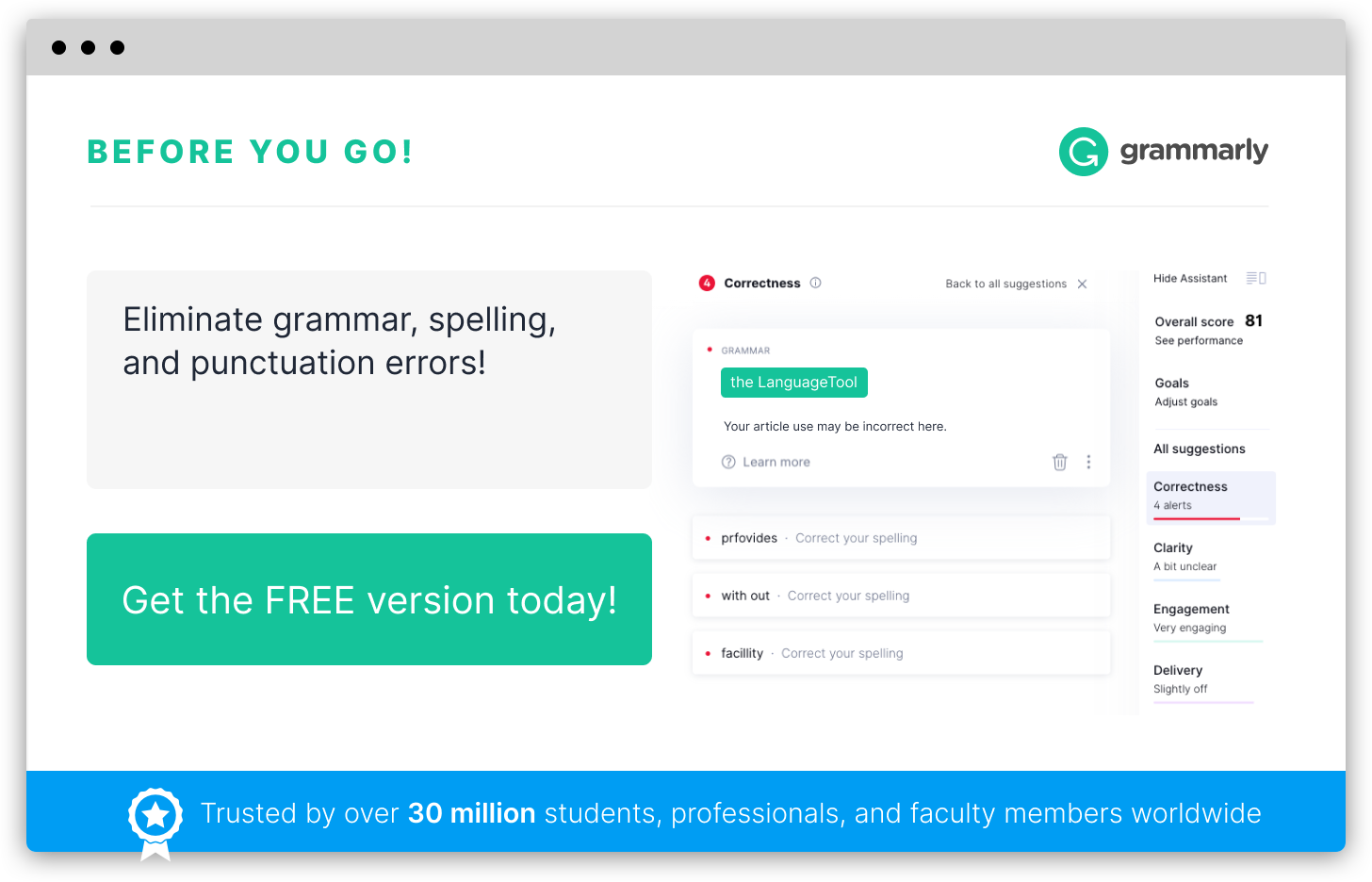
Spellcheck
Misspellings
Perhaps,
HUMAN_BRAIN
is a misspelling. Click
here
to search for this word in our misspelling dictionary.
Or did you mean:
human brain
X